Notatki
zjazd 4
Przepływ
energii w biosferze:
- Wszystkie substancje organiczne konieczne komórką do życia są produktami fotosyntezy,
- Zwierzęta zależą od roślin, które przekształcają energię świetlną w energię chemiczną,
- Energia chemiczna zawarta jest w cukrach i innych związkach organicznych.
Fotosynteza-
synteza związków organicznych z prostych substancji mineralnych
kosztem energii świetlej. Energia zostaje zmieniona na energię
chemiczną związków organicznych.
- Jako substraty służą CO2 pobierany z atmosfery przez liście i woda pobierana z gleby przez korzenie,
- Tlen uwalnia się jako jeden z końcowych produktów fotosyntezy,
- Wodór zostaje przeniesiony na CO2 powodując jego redukcję,
- Energii do procesu rozkładowi wody dostarcza światło pochłonięte przez chlorofil,
- Proces zachodzi w chloroplastach i jest bardziej skomplikowany niż przedstawione równanie
CO2+
2H2Oenergia świetlna CH2O+
O2
CH2O-
pierwotny produkt fotosyntezy, związek zredukowany do poziomu cukru.
Fotosynteza
składa się dwóch oddzielnych ale powiązanych ze sobą faz:
- Faza świetlna (fotochemiczna)- złożona z reakcji fotochemicznych wymagających światła. Polega na wykorzystaniu energii świetlnej do wytworzenia związków bogatych w energię: ATP i NADPH (są to siły asymilacyjne), zachodzi w błonach tylakoidów,
- Faza ciemna- nie wymaga światła, złożona jest z reakcji biochemicznych powodujących związanie i redukcję CO2, przy wykorzystaniu ATP i NADPH powstałych w fazie jasnej. Przebiega w Stromie chloroplastów.
Barwniki
fotosyntetyczne- występują w błonach tylakoidów i tworzą
kompleksy barwnikowo-lipidowo-białkowe, tzw. fotosystemy i
fotoukłady:
- U roślin występują dwa typy fotoskładów: fotoukład I i fotoukład II (inaczej określane jako PS I i PS II,
- Błona tylakoidy zawiera wiele tysięcy fotoukładów I i II,
- Pełnia rolę anten pochłaniających kwanty energii świetlnej (fotony) i przekazujących stan wzbudzenia do centrum reakcji fotochemicznej (tworzy je specjalna para cząsteczek chlorofilu a) ,
- W centrum reakcji fotochemicznej dochodzi do oddzielenia elektronu od wzbudzonej cząsteczki chlorofilu.
Barwniki
i ich występowanie:
- Chlorofil a – niebiesko-zielony, występuje we wszystkich roślinach zielonych,
- Chlorofil b- żółto-zielony, występuje we wszystkich roślinach zielonych z wyjątkiem pewnych glonów,
- Karoteny- pomarańczowo-czerwone, występuje we wszystkich roślinach zielonych, korzenie marchwi, niektóre owoce, nasiona,
- Ksantofile- żółta, żółta-pomarańczowa, występują w niektórych tkankach roślinnych zielonych i niezielonych.
Chlorofile-
są to główne barwniki fotosyntetyczne, nadające roślinom zieloną
barwę; absorbują światło w zakresie widzialnym, głównie w
niebieskiej i czerwonej części widma.
Budowa
chlorofilu:
- 5 – pierścieniowa pochodna porfiryny oraz dołączona do IV pierścienia pirolowego reszta 20- węglowego alkoholu- fitolu; hydrofobowy łańcuch fitolu umożliwia „zakotwiczenia” i odpowiednia położenie cząsteczki chlorofilu w błonie;
- Cztery pierścienie pirolowe oraz dodatkowy piąty pierścień niepirolowy tworzą układ następujących po sobie wiązań pojedynczych i podwójnych (tzw. układ wiązań sprzężonych) posiadających zdolność absorpcji promieniowania świetlnego,
- Centralne miejsce w pierścieniu porfirynowym zajmuje atom magnezu, połączony atomami azotu pierścieni pirolowych; pełnią ważną rolę w agregacji cząsteczek chlorofilu.
Światło
padające na chlorofil ulega częściowej absorpcji:
- Stan podstawowy- absorpcja kwantu promieniowania powoduje przejście chlorofilu do stanu wzbudzonego, czyli bardziej bogatego w energię; energia ta zostaje przekazana elektronom, które zostają przeniesione na orbity bardziej oddalone od jądra atomu,
- Energia wzbudzenia jest emitowana w postaci światła o długości fali dłuższej niż pochłonięte,
- Stan wzbudzony- stan nietrwały, cząsteczka dąży do pozbycia się nadwyżki energii.
Zawartość
barwników w liściach zależy od gatunku rośliny oraz warunków
wzrostu.
Zahamowanie
syntezy chlorofilu może być spowodowane:
- Brakiem światła (wyjątek stanowią niektóre mchy, glony, paprocie i szpilkowe),
- Deficyt azotu i magnezu,
- Brak żelaza i manganu w dostępnej formie,
- Zbyt niska lub zbyt wysoka temperatura,
- Odwodnienie tkanek,
- Infekcje bakteryjne grzybowe i wirusowe.
Przeciętnie
na 1 dm2 blaszki liściowej przypada 4-7 mg chlorofilu, przy czym
chlorofilu a jest zwykle 2-3 razy więcej niż chlorofilu b.
Karotenoidy:
- Określane są jako barwniki pomocnicze,
- Działają jak anteny przekazujące energię wzbudzenia na cząsteczkę chlorofilu a,
- Absorbują światło głównie niebiesko-fioletowe,
- Pełnią funkcję ochronną,
- Mogą przyjmować i rozpraszać nadmiar energii świetlnej i chronić w ten sposób błony tylakoidów oraz chlorofil przez fotoutlenianiem.
Światło
widzialne stanowi bardzo niewielką część ciągłego widma
(spektrum) promieniowania zwanego widmem elektromagnetycznym. Światło
widzialne jest mieszaniną fal o różnej długości: od 380 do 750
nm. Różnym zakresom światła widzialnego odpowiadają różne
barwy: fale najkrótsze- światło fioletowe, fale najdłuższe-
światło czerwone. Fale
krótsze dostarczają więcej energii a dłuższe mniej.
Schemat
funkcjonowania fotoukładów:
- W roślinach funkcjonują dwa typy fotoukładów (PSI= P-700 i PSII = P-680),
Faza
jasna fotosyntezy:
- Transport elektronów odbywa się z udziałem dwóch fotoukładów (PSI i PSII) oraz przenośników elektronów nie związanych z fotoukładami: PQA i PQB zawierające plastochinon, kompleks cytochromowy b6f, plastocyjanina, ferrdoksyna
- Właściwe reakcje polegają na absorpcji światła przez barwniki, wybiciu elektronów z centrów reakcji fotoukładów I i II, przekazaniu elektronów na pierwotne akceptory,
- Pozostałe reakcje przekazywania elektronów zachodzą zgodnie z gradientem potencjału oksydacyjno-redukcyjnego i mogą przebiegać również w ciemności,
- Podczas wędrówki elektronów następują stopniowe zmiany potencjałów Oksydo-redukcyjnych kolejnych przenośników,
- Wybicie elektronów z centrum reakcji PSII (stanowi je para cząsteczek chlorofilu a wykazującego maksimum absorpcji przy 680nm, stąd określenie P-680) powoduje powstanie kationu P-680+, który jest silnym utleniaczem i odciąga elektrony z wody. Prowadzi to do:
- Wydzielenia tlenu,
- Uwolnienia protonów,
- Powrotu centrum reakcji do stanu podstawowego (niewzbudzonego),
- Woda jest bezpośrednim donorem elektronów dla P-680+. Mechanizm rozkładu wody (fotoliza wody) na elektrony, protony i tlen jest złożony, ważną rolę odgrywają w nim atomy manganu. Kompleks enzymatyczny rozkładający wodę znajduję się na wewnętrznej powierzchni błony tylakoidy u zawiera 4 atomy manganu związane z białkiem o masie cząsteczkowej 33kDa,
- Transportowi elektronów z wody na NADP+ towarzyszy wytworzenie gradientu stężenia protonów w poprzek tylakoidu:
- Wzrost stężenia wewnątrz tylakoidu: uwalnianie protonów z rozkładu wody, Uwalnianie protonów z plastochinolu
- Obniżenie stężenia w Stromie: pro tonacja zredukowanego plastochinolu, redukcja NADP+
- Transport elektronów sprzężony jest z transportem protonów (H+) i wytworzeniem tzw. gradientu protonowego.
Notatki
zjazd 5
Mechanizm
fosforylacji fotosyntetyczne (fotofosforylacji):
- Mechanizm syntezy ATP w chloroplastach jest podobny do mechanizmu funkcjonującego w mitochondriach (fosforylacji oksydacyjnej),
- Siłą napędową syntezy ATP jest gradient protonów w poprzek błony tylakoidu tzn. między wnętrzem tylakoidu a stromą,
- Wnętrze tylakoidu staje się kwaśne, natomiast w Stromie dochodzi do alkalizacji,
- Gradient ten wynosi około 3,5 jednostek pH,
- W błony tylakoidów wbudowane są specjalne kompleksy białkowe (syntazy ATP) umożliwiające transport protonów z wnętrza tylakoidu do stromy i syntezę ATP.
Synataza
ATP z ADP i fosforanu nieorganicznego (Pi)
zachodzi z udziałem kompleksu syntazy ATP, zwanego czynnikiem
sprzęgającym. Przepływowi 3 protonów (H+) przez kompleks syntazy
ATP towarzyszy utworzenie 1 cząsteczki ATP.
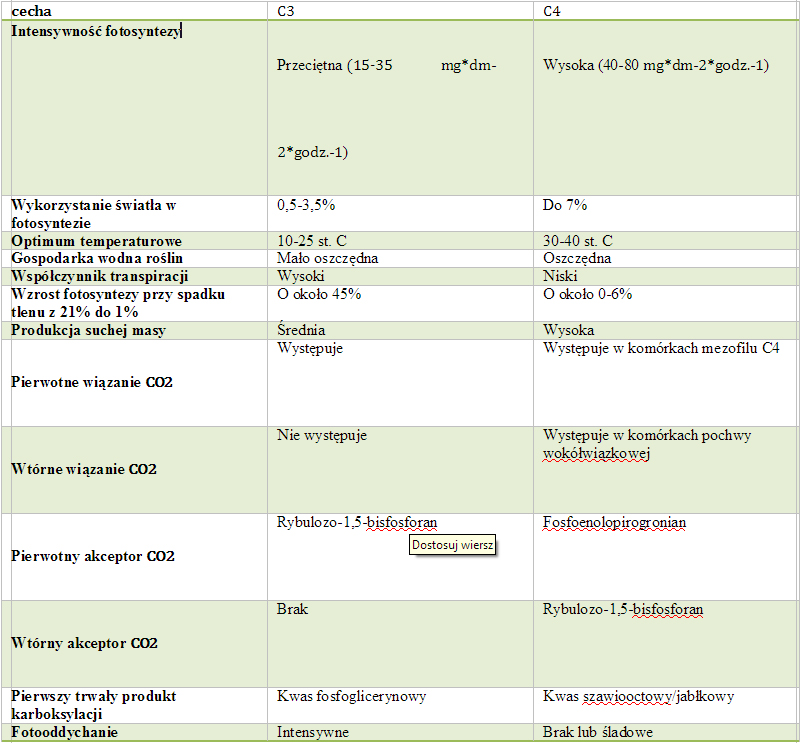
Faza
ciemna fotosyntezy (cykl Calvina-Bensona; cykl C3):
- Zachodzi w Stromie chloroplastów,
- Wyróżnia się trzy etapy:
- Karboksylacja- przyłączanie CO2 do cukru 5-cio węglowego-rybulozy; zachodzi przy udziale enzymy Rubisco (karboksylazy rybulozo-1,5-bisfosforanowej); powstają 2 cząsteczki kwasu 3-fosfoglicerynowego (PGA)
- Redukcja- redukcja kwasy fosfoglicerynowego (PGA) do aldehydu 3-fosfoglicerynowego (fosfotriozy); w procesie wykorzystywana jest tzw. siła asymilacyjna (ATP i NADPH) powstała w fazie jasnej fotosyntezy
- Regeneracja- seria skomplikowanych przemian umożliwiających odtworzenie akceptora CO2 czyli 1,5-bifosforybulozy; na początku 5 cząsteczek triozy ulega przekształceniu w trzy cząsteczki pentozy; metabolity pośrednie (cukry 4,5 i 6 węglowe) mogą być wykorzystywane do syntezy innych związków organicznych.
Fotorespiracja
(fotooddychanie):
- Proces kataboliczny zbliżony do oddychania,
- Polega na stymulowanym przez światło wydzielaniu CO2 związanym z pobieraniem tlenu,
- Nie generuje energii,
- Prowadzi do strat CO2 i zmniejszenia produktywności roślin i przyrostu biomasy (nawet do 50%),
- Przebiega w chloroplastach, peroksysomach i mitochondriach,
- Jest efektem specyficznej właściwości Rubisco , polegającej na tym, że poza podstawową funkcją wiązania CO2 może działać jako oksygeneza i wiązać tlen, co prowadzi do rozbicia 1,5-bisfosforybulozy,
- Zachodzi w roślinach typu C3.
Rubisco-
karboksylaza/oksygeneza rybulozo- 1,5- bis fosforanowa:
- Tlen i CO2współzawodniczą o to samo centrum aktywne enzymu,
- W normalnej atmosferze (0,036% CO2 i 21% O2) aktywność karboksyl azowa enzymu jest ok. 3-4 razy większa od aktywności oksygenazowej,
- Jeżeli stężenie obu gazów byłoby jednakowe karboksylacja zachodziłaby 80 razy szybciej niż reakcja utlenienia,
- Podwyższenie tem. Zwiększa temp procesu utleniania.
Cykl
Calvina: RuBP + CO2
-> 2PGA
Fotooddychanie:
RuBP + O2->
PGA + P-glikolan
- Utracony zostaje jeden atom węgla
- Uwalniany jest NH3
- Cykl zużywa 2ATP i 2,5 NADPH
Konsekwencje
fotooddychania:
- Powoduje stratę 25-40% węgla związanego w procesie fotosyntezy,
- Zmniejsza produktywność roślin nawet o 50%,
- Zapewnia funkcjonowanie fotosyntezy przy utrudnionym dopływie CO2,
- Odprowadza energię pochłoniętą przez barwniki fotosyntetyczne i chroni aparat fotosyntetyczny przed uszkodzeniami,
- Uaktywnia wewnątrzkomórkową wymianę i transport metabolitów.
Cykl
C4 (kwasów dwukarboksylowych):
- Niektóre rośliny wywodzące się ze strefy zwrotnikowej wykształciły dodatkowy mechanizm wiązanie CO2,
- Cykl C4 poprzedza reakcje cyklu Calvina,
- Wiąże się on z pewnymi cechami budowy anatomicznej liści,
- Został odkryty w 1965r. przez Kortschacka, Hatcha i Slacka.
Fizjologiczne
znaczenie podwójnej karboksylacja:
- Zwiększenie stężenia CO2 w komórkach pochwy około związkowej, a poprzez to wyeliminowanie strat węgla spowodowanych przez proces fotorespiracji,
- Większa wydajność fotosyntetyczna,
- Wysoki współczynnik wykorzystania azotu.
 Różnice metaboliczne:
Różnice metaboliczne: Wiązanie CO2 w roślinach typu CAM (niektóre gatunki rosnące w klimacie półpustynnym i pustynnym, głownie z rodzin: Crassulaceae, Cactaceae, Bromeliaceae):
Wiązanie CO2 w roślinach typu CAM (niektóre gatunki rosnące w klimacie półpustynnym i pustynnym, głownie z rodzin: Crassulaceae, Cactaceae, Bromeliaceae):- Cykl CAM- w czasie dnia, kiedy szparki są zamknięte jabłczan (4C) jest przenoszony z wakuoli do cytozolu, gdzie ulega dekarboksylacji. Uwolniony CO2 jest w chloroplastach wiązany w cyklu Calvina (Kleina;)

Brak komentarzy:
Prześlij komentarz